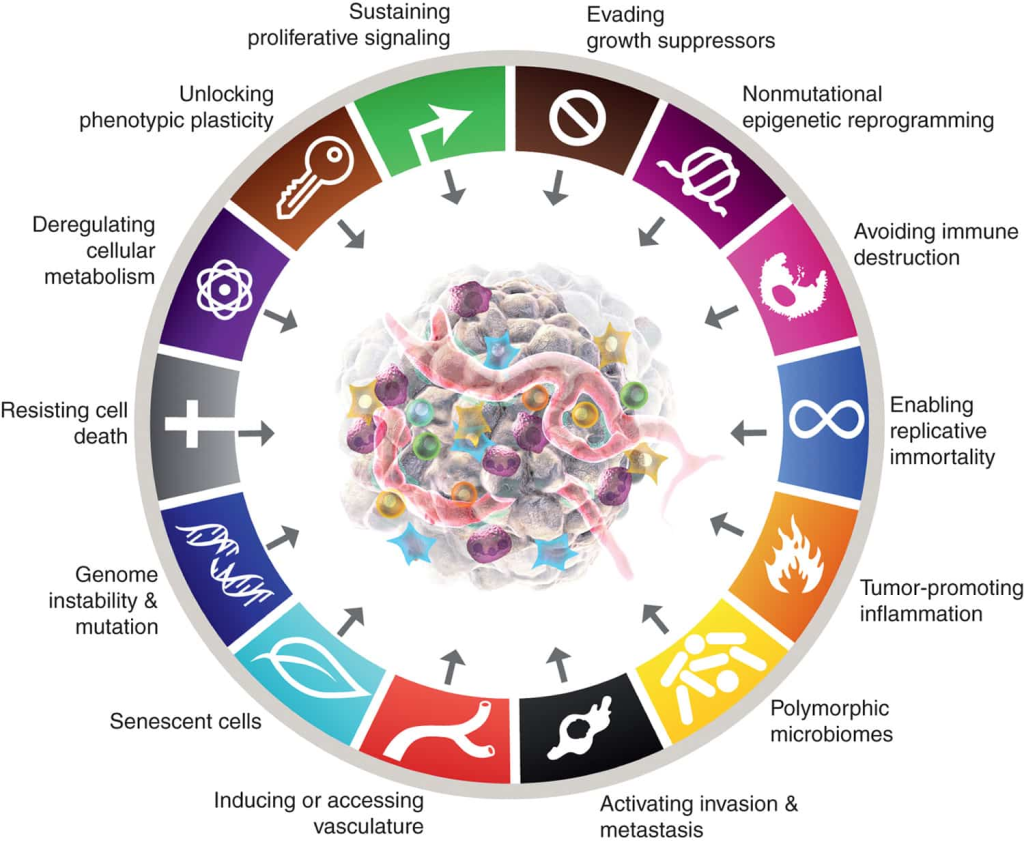
This is going to be a collection of metabolic curiosities in cancer metabolism:
Oxidised state through Fatty Acid Synthase
Prostate cancer cells stay more oxidised than their normal prostate cell counterparts. Fatty Acid Synthase works as an electron sink, where reduced electron carriers NADPH are oxidised back to NADP+
“The metabolic organization of both normal and malignant prostate cellular phenotypes involves some unusual and surprising features. In particular, both conditions exhibit ratios of NADH/NAD+ and NADPH/NADP+ charactersitic of high oxidative states despite a chronic shortage of O2 in both conditions. In this paper, we observe that, in prostate cancer cells, the oxidizing power of the fatty acid synthesis (FAS) pathway is so large that redox is stabilized more favorably (more oxidized) than in normal prostate cells.”
https://onlinelibrary.wiley.com/doi/10.1002/bies.10131
Desaturation of fatty acids – Cancers rely on PUFA / MUFA production to avoid apoptosis and probably to proliferate.
Speculation: Desaturated membrane lipids makes the membrane more water solluble. The taking up of water works as a proliferation signal. Cells that are more hydrophilic, tends to get bigger and divide more.
We have already made friends with SCD(1) on this blog in writing about how the SCD-1 enzyme on if the key drivers in mamallian hibernation and human obesity.
Blocking SCD 1, the enzyme that converts Saturated fats to monounsaturated fats, could even make the cell commit apoptosis again, probably changing the fatty acid composition in cardiolipin, it made the mitochondria able to release cytochrome C, and induce the apoptotic cascade.
“Using functional genomics, we identified stearoyl-CoA desaturase (SCD), an enzyme that controls synthesis of unsaturated fatty acids, as essential in breast and prostate cancer cells. SCD inhibition altered cellular lipid composition and impeded cell viability in the absence of exogenous lipids. SCD inhibition also altered cardiolipin composition, leading to the release of cytochrome C and induction of apoptosis. Furthermore, SCD was required for the generation of poly-unsaturated lipids in cancer cells grown in spheroid cultures, which resemble those found in tumour tissue. We also found that SCD mRNA and protein expression is elevated in human breast cancers and predicts poor survival in high-grade tumours. Finally, silencing of SCD in prostate orthografts efficiently blocked tumour growth and significantly increased animal survival.“
Seems that cancer takes advantage of the old hibernation adaptations.
https://cancerandmetabolism.biomedcentral.com/articles/10.1186/s40170-016-0146-8
—
Targeting metastasis-initiating cells through the fatty acid receptor CD36
Saturated fats or any fats from the diet seems to be important for metastasis initiating cancer cells.
“Palmitic acid or a high-fat diet specifically boosts the metastatic potential of CD36+ metastasis-initiating cells in a CD36-dependent manner. The use of neutralizing antibodies to block CD36 causes almost complete inhibition of metastasis in immunodeficient or immunocompetent orthotopic mouse models of human oral cancer, with no side effects. Clinically, the presence of CD36+ metastasis-initiating cells correlates with a poor prognosis for numerous types of carcinomas, and inhibition of CD36 also impairs metastasis, at least in human melanoma- and breast cancer-derived tumours. Together, our results indicate that metastasis-initiating cells particularly rely on dietary lipids to promote metastasis.“
https://pubmed.ncbi.nlm.nih.gov/27974793/
PUFA omega 3 induced expression of CD36
Induction of Cd36 expression elicited by fish oil PUFA in spontaneously hypertensive rats
https://pubmed.ncbi.nlm.nih.gov/16517147/
CD36 overexpression in CNS diseases. Promotes uptake of oxidised LDL.
CD36 in Alzheimer’s Disease: An Overview of Molecular Mechanisms
and Therapeutic Targeting
“To date, CD36 has been linked to several CNS pathologies, including chronic ischemia and stroke (Cho et al., 2005; Woo et al., 2016), familial forms of Parkinson’s disease (Abumrad and Moore, 2011), ALS (Stribl et al., 2014), frontotemporal dementia (Stribl et al., 2014), and AD (El Khoury et al., 2003; Sˇery´ et al., 2017). Most data are obtained from animal model studies. Clinical trials to address the relationship between CD36
and various neurodegenerative diseases are scarce, and those related to AD are discussed in the final sections of this review. CD36 upregulation favors atherosclerosis, chronic ischemia, and ischemic stroke (Silverstein, 2009). At the
cellular level, CD36 promotes atherosclerosis by uptake of oxidized LDL in macrophages, and thus, the formation of foam cells (Garcia-Bonilla et al., 2014). CD36 also induces heterodimeric binding of Toll-like receptors (TLRs), and the downstream activation of Nf-kB transcription factor for the production of pro-inflammatory cytokines, with deleterious effects in atherosclerosis and
stroke (Park, 2014).“
https://www.ibroneuroscience.org/article/S0306-4522(20)30719-3/pdf
Involved in metastasis
New role for CD36 in metastasis through fat intake
“It is well established that CD36 facilitates the uptake of long-chain fatty acids into adipocytes, the heart and the skeletal muscle. In this regard, adipose tissue expression of CD36 has been shown to correlate with body mass index suggesting a potential relevance in obesity (a key cardiovascular risk factor). Likewise, although cardiac CD36 mediates fatty acid uptake for energy production, enhanced CD36 expression in myocytes has been shown to favour heart lipotoxicity.“
High fat diet makes the tumor more agressive, neutralising CD36 neutralises metastatic potential.
“In the Nature article, the group of Salvador Aznar Benitah1 provides evidence of another function regulated by CD36 through fatty acid uptake, metastasis progression. The authors firstly demonstrated in human oral carcinomas that culprit cells behind oral cancers induce high levels of CD36 which, in turn, trigger a signaling cascade involved in the conversion of fatty acids to energy production (i.e. ATP). Interestingly, they evidence that CD36 metastatic potential is dependent on its lipid metabolism. As such, administration of a high-fat diet to mice induces the development of more aggressive oral cancers and more aggressive metastasis in a CD36-dependent manner. In counterpart, alteration of CD36 activity leads to intracellular accumulation of lipids and the consequent lipotoxic cell death (anti-metastatic effects). Based on these findings, the authors postulate that CD36+ cells may take advantage of this feature to take the amount of energy required to anchor and survive at sites distant from the primary tumour. The authors further demonstrate in vivo that infusion of these CD36+ cells to non-metastatic tumours induces metastasis, whereas, conversely, CD36 depletion by intraperitoneal administration of two different CD36-neutralizing antibodies not only prevents tumour metastasis but also largely regress those metastatic tumours that had already been developed. These anti-metastatic properties are observed without affecting primary tumour formation and without concomitant side effects”
https://academic.oup.com/cardiovascres/article/113/7/e16/3829657?login=false

